Embryonální vývoj, známý také jako zárodečný vývoj či embryogeneze, je fascinující proces, který se odehrává od prvního dělení zygoty až do stadia plodu (fetusu). Tento proces je klíčový pro vznik nového jedince a zahrnuje řadu komplexních změn na buněčné i molekulární úrovni.
Embryonální vývoj u živočichů
U obratlovců je embryonální období definováno jako období od prvního rozdělení zygoty (oplozeného vajíčka) až do doby, kdy se embryo vyvine v plod neboli fetus. Toto období zahrnuje klíčové procesy, jako je rýhování, vznik moruly a blastuly, diferenciace buněk, vývoj zárodečných listů a formování základů orgánových soustav.
Rané fáze embryonálního vývoje
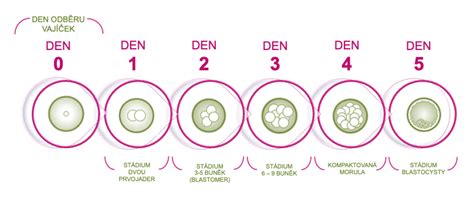
Vývoj embrya je komplexní proces, který i v době moderní lékařské vědy a pokročilé techniky nepřestává odborníky udivovat. Již v prvních dnech po oplodnění, v době, kdy nastávající matka ani netuší, že uvnitř jejího těla začíná nový život, probíhá jeden z nejdůležitějších procesů přírody. Během prvních 14 dnů urazí embryo neuvěřitelnou cestu. Z jedné jediné buňky se stává zárodek s propracovanou strukturou, připravený pro další vývoj.
Proces začíná spojením vajíčka a spermie, což vede ke vzniku zygoty. Tato nová buňka nese kompletní genetické informace od obou rodičů a je v ní zakódováno vše pro budoucí vývoj. Již pár hodin po oplodnění má zygota za úkol zreorganizovat DNA, vytvořit dvě jádra a spustit buněčné dělení. Přibližně 24 hodin od oplodnění se z jedné buňky stanou dvě nové. Druhá část buněk se podílí na vzniku podpůrných struktur, jako je placenta, žloutkový vak a plodové obaly.
Přibližně každých 10 hodin se embryo dále dělí, což představuje jedno z nejkritičtějších období jeho vývoje. Třetí a čtvrtý den vývoje dochází k dalšímu dělení na 8-16 buněk, které se k sobě začnou lepit. Embryo se začíná strukturovat a vzniká tzv. blastocysta. Uvnitř blastocysty se začne vyvíjet samotné miminko, zatímco z okolních buněk se bude vyvíjet placenta. V této době se embryo připravuje na uchycení v děložní stěně a zbavuje se ochranného obalu.
Buňky na povrchu embrya začínají prorůstat do výstelky dělohy. Právě v těchto dnech si přibližně 20-40 % žen může povšimnout slabšího krvácení, které je způsobeno uchycením embrya. Začíná také rychle stoupat hladina těhotenského hormonu hCG. Složitá síť propojuje embryo s krevním oběhem matky a embryo je již kompletně uchycené v děložní výstelce.
Embryo se rozvrství a vznikají dvě dutiny: vak, kde bude později plodová voda, a žloutkový vak zajišťující výživu do doby, než se plně vyvine placenta.
Procesy embryogeneze
Rýhování je vlastně mnoho po sobě jdoucích mitotických dělení. U běžných buněk bývá mezi dvěma děleními poměrně dlouhá interfáze, kdy dochází k přípravě na další dělení, ale u rýhujícího se vajíčka je interfáze velice redukovaná a neznatelná. Důležité je pro buňku pokaždé rychle replikovat (kopírovat) svou DNA - s přepisem DNA v RNA (tzv. transkripcí) se nezdržuje a veškerá buněčná RNA pochází ještě od matky.
Rýhování může být dvojího druhu: buď úplné (totální, holoblastické), nebo částečné (parciální, meroblastické). Úplné rýhování probíhá u vajíček s nízkým nebo žádným obsahem žloutku (např. savci, měkkýši nebo hlístice).
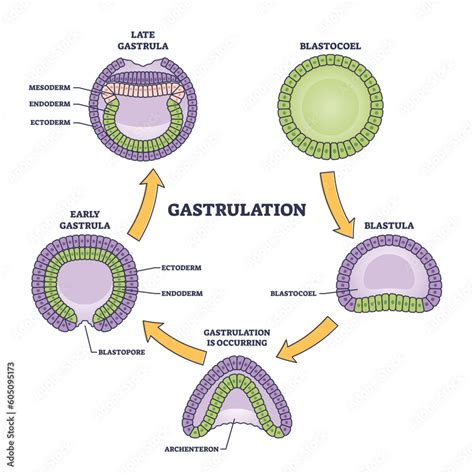
Blastula je rané embryonální stadium, které má kulovitý tvar a uvnitř obsahuje dutinu, blastocoel. V klasickém schématu embryonálního vývoje vzniká v procesu blastulace (blastogeneze) z moruly a vyvíjí se z ní gastrula. Podoba blastuly závisí na typu rýhování, jímž dělící se zárodek procházel. V některých případech dokonce blastula vůbec nemusí mít zřetelný blastocoel (je plná). Blastula u savců se nazývá blastocysta. Ta se pohybuje volně v děloze, vyživuje se jejím sekretem, ale nakonec (u člověka během 6. týdne) se uchytí v děložní stěně.
V průběhu rané embryogeneze začínají obvykle vznikat zárodečné listy. To platí pro všechny pokročilé živočichy, zahrnované obvykle do Eumetazoa. Podle toho, zda vznikají listy dva nebo tři, rozlišujeme živočichy dvojlisté a trojlisté. Zárodečné listy vznikají v době, kdy se blastula mění na gastrulu. Tento proces se nazývá gastrulace. Gastrula má nakonec již dvě vrstvy buněk a obsahuje dutinu (tzv. prvostřevo), která se otvírá do okolního světa otvorem zvaným blastoporus.
Podle nejčastěji uváděného schématu gastrulace se buňky na jednom konci kulovité blastuly začnou pohybovat směrem dovnitř dutiny, čímž vytváří jakýsi polokulovitý dutý útvar (přirovnávaný k prokopnutému míči). Migrující buňky posléze téměř přilnou k těm na opačné straně a v místě, kde zprvu začala gastrulace, jsou teď prvotní ústa (tedy budoucí ústa prvoústých). Gastrulace však probíhá u různých organizmů různě a zrovna způsob gastrulace u člověka je poměrně specifický. Závisí totiž na typu blastuly, z níž gastrula vzniká. Z coeloblastuly vzniká coelogastrula přibližně tak, jak bylo právě popsáno. Jiná situace nastává u blastul, které obsahují velké množství žloutku nebo například vůbec nejsou duté.
Klíčem ke vzniku notochordu (struny hřbetní) u lidského embrya je proces gastrulace: nořící se epiblastové buňky vytváří tímto způsobem entodermální a mezodermální tkáně. Jednou z nich je i notochord. Část buněk vnikajících primitivní brázdičkou do spodní části embrya se stáčí směrem k tzv. prechordální ploténce, o tu se zastaví a vytvoří hlavový úsek notochordu. Následně sice nakrátko zaniká, ale později se opět zvýrazňuje a tvoří pevnou osu vedoucí od Hensenova uzlu (na jednom konci brázdičky) až po prechordální ploténku.
Formování orgánových soustav
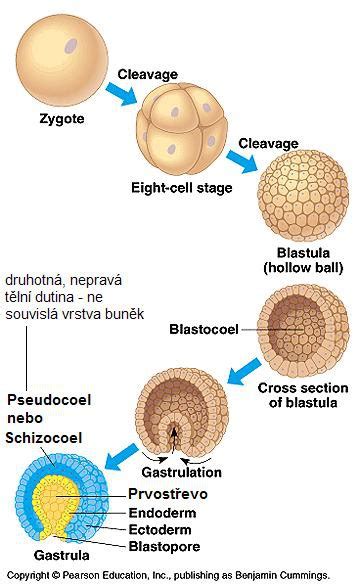
Embryonální stadium, v němž vzniká neurální trubice, se nazývá neurula - vlastně je to však pozdní gastrula. Spouštěčem vzniku neurální trubice je vznik struny hřbetní (chorda dorsalis). V této souvislosti se říká, že dochází k neurální indukci. Ektoderm ležící nad chordou začne díky tomu tlustnout a formuje se nejprve tzv. neurální ploténka, jejíž tkáň se označuje jako neuroektoderm. Tím začíná celý proces neurulace.
Následně se buňky nervové ploténky vchlipují, okraje ektodermu nad ním („neurální valy“) se začnou vzájemně přibližovat a nakonec splývají, čímž vzniká pod ektodermem dutá neurální trubice. Neurulace probíhá poněkud odlišně v hlavové části, než ve zbytku těla, velkou roli tam totiž hraje tzv. kaudální morfogenetický systém. Proces neurulace je poté víceméně u konce, vzniklá nervová trubice má uvnitř tzv. neurocoel.
V pozdějších fázích vývoje embrya však dojde k velkým tvarovým změnám nervové trubice, při nichž vznikají komplikovanější struktury, jako je mícha nebo mozek. S procesem neurulace souvisí také vznik neurální lišty, což je populace buněk, které vzniknou po odškrcení nervové trubice od zbylého ektodermu.
Tělní segmentace je ze všech živočichů asi nejrozvinutější u členovců a podrobně zkoumána byla u octomilky („mouchy“ rodu Drosophila). U strunatců, jako je například člověk, však dochází rovněž ke vzniku segmentů, které se označují somity či také primitivní segmenty. Jsou to párové oblasti mezodermu, které se vytváří v raných fázích vývoje a leží na obou stranách podél struny hřbetní. Obecně platí, že se odštěpují jako váčky z coelomu. Posléze vyplňují prostor kolem neurální trubice a nakonec vrůstají i do bočních mezer mezi střevem a stěnou těla. U kopinatců je vývoj poněkud odlišný než například u ryb, ale obecně ze somitů vznikají zejména svaly, někdy však i podkožní vazivo, vylučovací soustava atd.
Žaberní oblouky (arcus branchiales) jsou přepážky, které oddělují jednotlivé žaberní štěrbiny. Vyskytují se v embryonálním stadiu většiny obratlovců. Všichni obratlovci se totiž vyvinuli z předků ve vodě. Ti měli zřejmě 9 párů žaberních štěrbin a oblouků. Kruhoústí (primitivní obratlovci) mají tento stav do velké míry zachovaný, ale již u čelistnatých obratlovců došlo k velkým stavebním změnám, díky nimž vznikly čelisti. Například u ryb jsou žaberní oblouky sloužící k výměně plynů pouze čtyři, pátý bývá přeměněn na požerákové zuby. Tyto rybí žaberní oblouky se samozřejmě u suchozemských obratlovců ve své původní funkci nezachovaly; přesto se během embryonálního stadia zakládají a vytváří neuvěřitelné množství struktur v lidské hlavě i krku.
Embryo v rostlinné říši
V botanice je embryo součástí semene. Semeno obsahuje ještě zásobní látky, ze kterých může embryo čerpat energii a prvotní stavební materiál při klíčení a vývoji klíční rostlinky.
Resyntéza řepky olejky
Řepka olejka (Brassica napus L. ssp. oleifera) patří mezi nejdůležitější olejniny. Z hlediska globální produkce oleje je na třetím místě za palmou olejnou a sójou, v Evropě jde o olejninu klíčovou. Intenzivním šlechtěním řepky dochází k neustálému zlepšování výnosu semene a dalších agronomických vlastností.
Šlechtění je v současnosti zaměřeno nejen na tvorbu vysoce výnosných odrůd, ale také na specifické složení mastných kyselin v oleji, zvýšenou rezistenci vůči významným chorobám a abiotickým stresům. Nabývá na důležitosti i šlechtění na adaptabilitu ke specifickým půdně-klimatickým podmínkám. Jedním z nejdůležitějších předpokladů tvorby výkonných odrůd se ukazuje dostatečná genetická variabilita výchozích materiálů, protože dlouhodobé šlechtění na relativně úzký okruh vlastností u řepky postupně vede k zužování genetické diverzity šlechtitelských materiálů.
Řepka olejka vznikla s největší pravděpodobností spontánním mezidruhovým křížením brukve řepáku (řepice, B. rapa) a zelné brukve (B. oleracea) nejspíše již v prehistorické době. Nelze ale mluvit o zkřížení v klasickém slova smyslu, které závisí na spárování příslušných chromozómů od obou rodičů. Chromozómy řepice a brukve se vzájemně nepárují, výsledkem oplození je tak zygota řepky se sadou chromozómů, odpovídající prostému součtu chromozómů obou pohlavních buněk (gamet) řepice a brukve, tj. n = 10 + 9 = 19. Protože každý chromozóm je v této zygotě zastoupen jen v jednom exempláři, jde o hybrid pouze s polovičním (haploidním) počtem chromozómů, podobně jako tomu je u gamet rodičů. Pravděpodobně již v počátečních fázích vývoje haploidního embrya však může dojít k samovolnému zdvojení chromozómové sady, čímž embryo získá normální (diploidní) počet chromozómů 2n = 2 x 19 = 38.
Protože přírodní populace řepky neexistují, dá se předpokládat, že spontánní vznik řepky výše uvedeným způsobem byl unikátní jev. Z tohoto důvodu také není možné pro obohacení genofondu - na rozdíl od dalších významných plodin - použít „plané“ řepky z přírodních sběrů. I proto byly v minulosti učiněny pokusy o umělé znovuvytvoření (tzv. resyntézu) řepky hybridizací brukve a řepice, nejprve v polních podmínkách.
Zástupci výchozích komponent pro resyntézu (B. rapa - řepice, vodnice a B. oleracea - kadeřávek, zelí, kedluben, kapusta, květák, brokolice apod.) se totiž vyznačují širokou variabilitou z hlediska morfologie i agronomických charakteristik a představují perspektivní zdroj heteróze a dalších znaků. I když byly experimenty úspěšné, počet takto získaných, životaschopných a fertilních řepk byl velmi nízký. Proces je komplikován jak nutností zdvojit chromozómovou sadu haploidní rostliny, tak i nefunkčností endospermu vznikajícího semene, který není schopen zárodek vyživovat a embryo odumírá již v počátečních fázích vývoje. Značný posun přineslo až využití in vitro biotechnologií.
Biotechnologické postupy v pěstování řepky
Postup křížení
Výchozí otcovské a mateřské rostliny brukve a řepice jsou před začátkem kvetení umístěny do podmínek s řízenou teplotou, světelnou intenzitou a fotoperiodou (fotoperioda 16/8 h, teplota den/noc 18/14 °C, intenzita osvětlení 84 μmol/m2/s), aby byl zajištěn optimální vývoj pylových zrn, resp. minimalizován vliv prostředí na rychlost a rovnoměrnost vývoje embryí v šešulích. Otcovské rostliny (B. oleracea) jsou před začátkem kvetení jednotlivě izolovány např. ve vacích z netkané textilie, aby nedošlo ke kontaminaci pylu jejích květů pylem z dalších otcovských nebo mateřských komponent. Před započetím křížení je třeba odstranit květy mateřské rostliny, aby se zabránilo nežádoucímu sprášení pylem z okolních květů. Po otevření poupat mateřské rostliny (B. rapa) a odstranění všech šesti prašníků pinzetou následuje přenesení pylu z květů otcovské rostliny (např. smotkem buničité vaty) na blizny mateřské rostliny. Aby byla vyloučena možnost nežádoucího opylení, je třeba dbát na desinfekci rukou a nářadí, např. 70% roztokem etanolu. Po opylení jsou jednotlivá květenství izolována sáčkem z celofánu, ten fixován kancelářskými sponkami ke květnímu stvolu nebo nejbližšímu listu a křížení označeno identifikačním štítkem (datum křížení, identifikace otcovské rostliny a počet opylených poupat).
Postup tzv. embryo rescue
Jedenáctý den po opylení jsou jednotlivé šešule i se stopkou odstřiženy z mateřské rostliny, umístěny do kádinky a přeneseny do laboratoře v cestovní chladničce. V laminárním boxu proběhne povrchová sterilizace šešulí 70% etanolem po dobu 2 minut, následovaná 20% roztokem SAVA po dobu 20 minut. Po opláchnutí šešulí sterilní chlazenou destilovanou vodou (3 cykly po 5 minutách) jsou šešule na filtračním papíru pod binokulární lupou podélně naříznuty ostrým skalpelem, nezralá vajíčka šetrně vyjmuta jemnou pinzetou nebo preparační jehlou a přenesena na povrch kultivačního média DM v Erlenmeyerových baňkách. Po uzavření baněk jsou nádobky přeneseny do kultivační místnosti s řízeným režimem (fotoperioda den/noc 16/8 h, světelná intenzita 250 μmol/m2/s, 25 °C). První embrya se objevují zhruba 20-30 dní po izolaci. Zelená embrya o délce 2-3 mm jsou přenášena na čerstvé DM médium.
Výše uvedený postup se nazývá embryo rescue, protože embrya jsou „zachraňována“ na umělých živných půdách, nahrazujících nefunkční endosperm semene. Úspěšnost embryo rescue závisí jak na šetrnosti manipulace s izolovanými nezralými vajíčky, tak na kombinaci výchozích rodičů (tj. genotypu). Pro získání alespoň jednoho embrya je obvykle třeba kultivovat nezralá vajíčka z 1-3 šešulí.
Postup regenerace rostlin a klonování
Jakmile embrya dosáhnou délky cca 8-10 mm, jejich děložní lístky jsou z 1/2 až 2/3 odříznuty pro podpoření vývoje vrcholového (apikálního) meristému. Následuje přenos ořezaných embryí na médium RM a snížení teploty v kultivační místnosti z 25°C na 19°C. Rostliny s 1-2 pravými lístky jsou přemístěny na čerstvé RM médium, rostliny ve fázi 3-5 pravých lístků pak na MS médium.
Protože všechny takto regenerované rostliny mají poloviční počet chromozómů, je u nich třeba provést již zmíněné zdvojení chromozómové sady. Jelikož se tento proces neobejde bez citelných ztrát rostlinného materiálu, je doporučováno jednotlivé rostliny ještě in vitro podmínkách namnožit klonováním z nodálních segmentů. Obvyklý postup je příčné rozdělení stonků rostlin v místech nad a pod spícím očkem s následnou regenerací rostlin z oček segmentů na MS médiu. Jako dostatečný počet pro dopěstování alespoň jedné životaschopné rostliny se zdvojenou sadou chromozómů do fáze kvetení se uvažuje cca 10 ks naklonovaných jedinců.
Postup zdvojení chromozómové sádky a dopěstování rostlin
Rostliny jsou vyjmuty z kultivačních nádob, opláchnuty pod vlažnou tekoucí vodou pro odstranění zbytků agarového média. Následuje krátké osušení regenerantů na filtračním papíru, ponoření celých rostlinek do 0,05% roztoku kolchicinu, zakrytí nádoby potravinářskou fólií a umístění do kultivační místnosti do 25°C na 24 hod. Poté jsou rostliny opláchnuty pod mírným proudem vlažné vodovodní vody, ponořeny do 0,15 % roztoku Previcuru na 20 minut, vysázeny do květníků s výsevním substrátem (nebo perlitem) a umístěny do přepravky. Aby se zabránilo vadnutí rostlin, květníky v přepravce musí být po dobu 7-10 dní zakryty perforovanou fólií.
Následuje dopěstování rostlin konvenčními postupy (přesazování do větších nádob, jarovizace - pokud alespoň jeden z rodičů byl ozimého typu, průběžná izolace celých rostlin nebo jednotlivých květenství před počátkem kvetení izolačními sáčky, odstraňování sterilních rostlin bez prášivého pylu a u rostlin s neschopností samoopylení v květu - obvykle z důvodu autoinkompatibility - ruční opylování v poupěti. Rostliny se v průběhu dopěstování pravidelně přihnojují a kontroluje jejich zdravotní stav, příp. se ošetřují proti chorobám a škůdcům.
K největším ztrátám obvykle dochází ve fázi převodu ze sterilních do nesterilních podmínek, protože jsou rostliny od počátku navyklé na prostředí 100% vzdušné vlhkosti bez přítomnosti patogenů a udržované v ideálních světelných, teplotních a výživových podmínkách. Ztráty lze v případě dobré kondice in vitro rostlin, šetrnou manipulací před/po zdvojování sádky a při vysazování, a umístěním do vhodných podmínek udržet pod 20 %. Při zanedbání i jedné z výše uvedených náležitostí počet rostlin nezřídka klesne na 0-30 % výchozího počtu.
Úspěšnost zdvojení chromozómové sady haploidů (tzv. dihaploidizace) je obvykle 30-60 %. Limitující okolností pro získání fertilních rostlin je však vhodná kombinace výchozích rodičů. Velmi často se stává, že z důvodu nekompatibility genotypů jsou všechny rostliny z dané kombinace buď sterilní, nebo mají poruchy fertility, nesouvisející s ploidií (neprašivý pyl, zakrnělé blizny, nízká nebo žádná násada semen apod.).
I když využití resyntetizovaných genotypů k přímé tvorbě odrůd řepky olejky není možné zejména z důvodu horší produktivity a dalších důležitých agronomických vlastností, v našich experimentech i na pracovišti šlechtitelů již bylo ověřeno, že požadované vlastnosti lze přenést křížením z resyntetizovaných řepk do konvenčních šlechtitelských materiálů. Unikátnost resyntézy řepky spočívá především v tom, že lze vytvořit zcela nové genotypy s vlastnostmi, které v současnosti nemusí být k dispozici v dosažitelném genofondu (odrůdy, šlechtitelské materiály, položky genové banky). Může se jednat např. o odolnost k suchu a chorobám, rychlejší růst v podzimním období, specifická kvalita oleje, netradiční habitus rostlin apod. Biotechnologické postupy, zavedené a optimalizované na našem pracovišti lze tak úspěšně využít k rozšíření diverzity genofondu této strategické plodiny.
Somatic Embryogenesis Explained 🌱 | Plant Tissue Culture Basics
tags: #deleni #rostlinneho #embrya